Bibliography Background About KRIS
Water Temperature and Gulf of Maine Atlantic Salmon
Sensitivity by Life Stage
Effects of Disease and Competition
Factors That Influence Water Temperature
Atlantic salmon have higher tolerance to warm water temperatures than Pacific salmon species (McCullough, 1999) and their closer cousins in the genus Salar, the Arctic charr and brown trout (Jensen et al., 1989). Warm temperatures can reduce fecundity, decrease egg survival, retard growth of fry and smolts, reduce rearing densities, increase susceptibility to disease and decrease the ability of young salmon and trout to compete with other species for food and to avoid predation (McCullough, 1999). The Maine "Atlantic Salmon Technical Advisory Committee (Dill et al., 2002) expressed concern about the potential impacts of increased water temperature on Gulf of Maine Atlantic salmon stocks:
"Maine rivers lie near the southern extent of the Atlantic salmon’s range in, and are vulnerable to elevated water temperature regimes. Contributing factors include improper or unregulated land use practices, impoundment of free-flowing reaches (where applicable), discharge of industrial processing or cooling water, and broad climatic changes. Relatively minor increases in water temperature may diminish habitat suitability for adult and juvenile salmon, lead to sub-lethal or lethal physiological responses, and have a profound negative effect on the production potential of a river or stream."
Cold water temperatures in Gulf of Maine rivers during winter may also limit Atlantic salmon growth and survival because temperatures dip below 4° C, which is limiting to growth in all life history phases (Armstrong et al., 2004). Formation of anchor ice can also be a major problem for salmonids during periods of severe cold, although juveniles tend to move to deeper areas durung winter (Hendry and Cragg-Hine, 2003).
|
 |
|
| This photo shows the West Branch Sheepscot River at Weeks Mills Road in Fall. Click to enlarge photograph. | Same location with ice encroaching. Photos by Melissa Halsted. Click to enlarge. | Surface of the West Branch Sheepscot River nearly ice covered. Click to enlarge photo. |


Elliot and Hurley (1997) defined the lower and upper temperature limits for growth of Atlantic salmon as 6.0° and 22.5° Celsius (C), and 15.9° C as the optimum temperature. Crisp (1993) indicated that the species shows signs of stress at approximately 22° C and that upper lethal limits were between 25° and 28° C. Swansburg et al. (2002) and the Maine Atlantic Salmon TAC (Dill et al., 2002) expressed concern over possible changes in Canadian and Maine rivers, respectively, related to global warming, with models suggesting reduced future growth rates of Atlantic salmon juveniles. Table 1 summarizes temperature tolerance values by life history stage for Atlantic salmon.
Table 1. Atlantic salmon water temperature tolerance in degrees Celsius by life history stage.
| Life Stage | Optimum Range | Minimum | Maximum | Literature Cited |
| Spawning | 5-8° C |
4.0° C |
10-12° C |
DeCola (1970), Danie et al. (1984), Beall and Marty (1983) |
| Egg/Alevin | 4-7.2° |
0.5° |
12° |
DeCola (1970), Peterson et al. (1977), Elliot et al. (1998) |
| Early Fry | 8-19° |
0.5° |
23.5-27.7° |
Danie et al. (1984), Jensen et al. (1991) |
| Parr | ||||
| Feeding | 15-19° |
3.8° |
22.5° |
DeCola (1970, Elson (1975), Danie et al. (1984), Elliott (1991) |
| Survival | 0.5-20° |
0° |
27-32° |
Garside (1973), Elliot (1991), Elliott and Elliott (1995) |
| Smolt (Migrating) | 7-14.3° |
5° |
19° |
Bakshitansky et al. (1979), LaBar et al. (1978), Jonnson and Rudd-Hansen (1985), Duston et al. (1991), Shepard (1991) |
| Adult (Migrating) | 14-20° |
8° |
23° |
Elson (1969), Danie et al. (1984), Shepard, S.L. (1995) |
 Eggs are the most temperature sensitive Atlantic salmon life history stage (Danie et al., 1984). While they thrive at temperatures as high as 7° C, when temperatures increase to 12° C there is a substantial increase in fungus, which may lead to mortality (DeCola, 1975). Elliot et al. (1998) found that mortality of eggs increases as water temperatures drop below 4 C and Peterson et al. (1977) noted that very low survival when temperatures dropped below 2° C. The gestation of Atlantic salmon eggs and alevin are substantially extended when low water temperatures prevail, and fry may not emerge from the nest for over 180 days after spawning (Baum, 1997). In order to insure that eggs are at least in an eyed stage when winter water temperatures drop, spawn timing of Atlantic salmon does not extend later than mid-November.
Eggs are the most temperature sensitive Atlantic salmon life history stage (Danie et al., 1984). While they thrive at temperatures as high as 7° C, when temperatures increase to 12° C there is a substantial increase in fungus, which may lead to mortality (DeCola, 1975). Elliot et al. (1998) found that mortality of eggs increases as water temperatures drop below 4 C and Peterson et al. (1977) noted that very low survival when temperatures dropped below 2° C. The gestation of Atlantic salmon eggs and alevin are substantially extended when low water temperatures prevail, and fry may not emerge from the nest for over 180 days after spawning (Baum, 1997). In order to insure that eggs are at least in an eyed stage when winter water temperatures drop, spawn timing of Atlantic salmon does not extend later than mid-November.
 During the parr life stage Atlantic salmon may be limited by cold water temperatures as well as elevated ones. "The growing season for Atlantic salmon has been defined as the number of days during which the air temperature exceeds 5.6° C (Power, 1981) or the water temperature exceeds 7° C (Symons, 1979)" (as cited in Armstrong et al., 2004). Heggenes (1990) noted that Atlantic salmon parr burrowed in the substrate of streams when water temperatures were at winter lows, most likely to come in contact with warmer ground water. Anchor ice may form in severely cold years freezing streams solid in some reaches.
During the parr life stage Atlantic salmon may be limited by cold water temperatures as well as elevated ones. "The growing season for Atlantic salmon has been defined as the number of days during which the air temperature exceeds 5.6° C (Power, 1981) or the water temperature exceeds 7° C (Symons, 1979)" (as cited in Armstrong et al., 2004). Heggenes (1990) noted that Atlantic salmon parr burrowed in the substrate of streams when water temperatures were at winter lows, most likely to come in contact with warmer ground water. Anchor ice may form in severely cold years freezing streams solid in some reaches.
Interestingly, Jensen (1990) found that Atlantic salmon parr grew at a faster rate at a given water temperature in spring than at the same temperature in fall, when streams were cooling. Bult et al. (1999) noted that juvenile Atlantic salmon selected feeding areas with slower currents when water temperatures were elevated and Metcalfe et al. (1997) postulated that such movement and loss of territorial behavior would reduce growth. Hendry and Cragg-Hine (2003) described seasonal behavior changes driven by temperature: "Juvenile salmon leave the riffles in autumn as temperatures decrease, and spend the colder months in the deeper pools, reappearing in shallower water in spring when temperatures reach 6 to 7°C."
Lethal temperatures for Atlantic salmon derived in the laboratory may vary depending on periods of acclimation (see Table 2). When water temperatures are raised rapidly, fish may survive briefly at temperatures of 32° C or higher, but the lethal level drops closer to 27° C when water temperatures are raised more slowly (Elliot and Elliot, 1995). Gradual increases are more similar to conditions in nature and some laboratory experiments may not reflect survival in streams. Fish in the wild must forage for food and avoid predation, while in laboratory environments the fish are fed and protected from predators. Stress may also occur at lower temperatures in the wild as the fish must cope with all the variables of its environment.
Table 2. Heating rates or durations and incipient lethal water temperatures for juvenile Atlantic salmon fry and parr.
Age Class |
Acclimation Temperature | Heating Rate | Incipient Lethal Temperature | Literature |
| 0+ and 1+ | 20° C | 0.1° C/hr. 1° C/hr. 18° C/hr. |
27.74° C (±1.20) 32.63° C (±0.38) 32.83° C (±0.83) |
Elliott and Elliott (1995) as cited in McCullough (1999) |
| 0+ and 1+ | NA | NA | 27.8° C at 7 days
29.5° C at 1000 minutes 31.1° C at 100 minutes 32.9° C at 10 minutes |
Elliott (1991) as cited in Dill et al. (2002) |
| 2-3 month old fry | 17° C | .04-.08° C/hr. | 28.7-29.2°C | Grande and Anderson (1991) as cited in McCullough (1999) |
| 3-4 month old fry | 17° C | 0.08° C/hr. | 29.2°C | Grande and Anderson (1991) as cited in McCullough (1999) |
 Danie et al. (1984) defined Atlantic salmon spawning temperatures as 4.4° and 10° C, while Beall and Marty (1983) found that fish would spawn in water as warm as 12° C, but not in warmer conditions. Adult salmon sometimes die when temperatures reach 26-27° C for several days and adults will not migrate upstream in water temperatures over 23° C (Garside, 1973; DeCola, 1970; Danie et al., 1984; Hawkins, 1989; Shepard, 1995). By contrast, low water temperatures may impede adult spawning migrations in places like northern Canada where entry of Atlantic salmon into rivers in spring is keyed by when river temperatures warm (Mills, 1991).
Danie et al. (1984) defined Atlantic salmon spawning temperatures as 4.4° and 10° C, while Beall and Marty (1983) found that fish would spawn in water as warm as 12° C, but not in warmer conditions. Adult salmon sometimes die when temperatures reach 26-27° C for several days and adults will not migrate upstream in water temperatures over 23° C (Garside, 1973; DeCola, 1970; Danie et al., 1984; Hawkins, 1989; Shepard, 1995). By contrast, low water temperatures may impede adult spawning migrations in places like northern Canada where entry of Atlantic salmon into rivers in spring is keyed by when river temperatures warm (Mills, 1991).
Effects of Disease and Competition
Substantial research demonstrates that many fish diseases become more virulent at temperatures over 15.6º C (McCullough, 1999). Some diseases, like infectious hematopoietic necrosis (IHN) that are present in Maine, however, are triggered by low water temperatures. Fish disease organisms are always present in the water, but as salmonid species become stressed by higher temperatures their resistance drops and the likelihood of an epidemic increases (Gullien, 2003). "Also, diseased fish probably are more susceptible to predation and less able to perform essential functions, such as feeding, swimming, and defending territories" (McCullough, 1999). Hatchery reared salmonids are more susceptible to disease than wild fish because of high fish rearing densities (Baum, 1997).
Baum (1997) noted that disease outbreaks in wild Atlantic salmon in Maine rivers are not common. Danie et al. (1984), however, noted that "temperatures of 20-27° C reduce resistance to disease and therefore are indirectly lethal" to Atlantic salmon. Common fish diseases in Maine such as Ich (Ichthyophthirius multifiliis) and Columnaris (Flavobacterium columnare) are known to pose greater risk to salmonids when water temperatures are elevated. Dickerson and Dawe (1995) noted that fish were more susceptible to Ich when stressed and when water temperatures were elevated, although incidence increased with crowding. Guillen (2003) noted that Columnaris also becomes more communicable during periods of elevated water temperature and low flow and that the two diseases may also act together on stressed fish as resistance drops after initial infection.
While salmonids may survive warm water temperatures in laboratory conditions, competition with warm water tolerant species in the wild may make survival problematic. Competitive interactions can reduce the ability of salmonids to maintain feeding stations and grow in streams with warmer temperatures (McCullough, 1999). There are numerous warm water adapted species that have been introduced as game fish to Gulf of Maine rivers (Meister, 1982). The performance of these fish improves with increasing water temperatures, while Atlantic salmon move away from preferred habitats and become less competitive and likely more subject to predation.
Factors That Influence Water Temperature
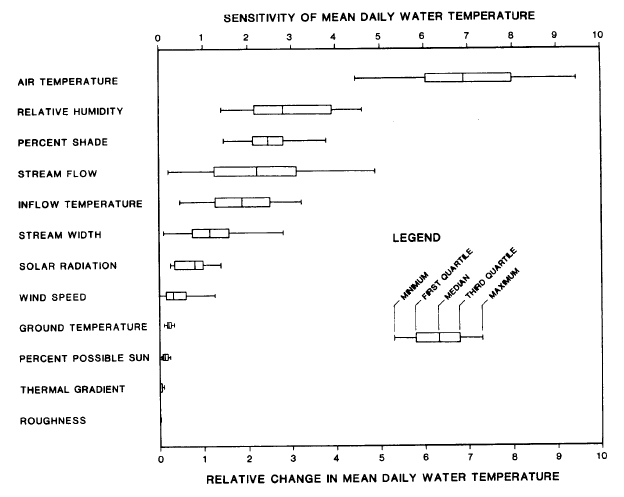 Bartholow (1989) found that air temperature above the stream surface was the greatest factor in increasing water temperatures followed in importance by relative humidity and shade, respectively (see graph at left). Bartholow's (1989) work was based on field data from hundreds of locations throughout the West used to develop the SNTEMP stream temperature model. While many previous works considered direct solar radiation to be the dominant mechanism for warming streams (Brown, 1980 as cited in Spence et al., 1998), most of the recent scientific literature considers air temperature over the stream to be the most influential factor (see quote from Essig, 1998 ). Poole and Berman (2000) also recognize the relationship between increasing air flow over the stream and water temperature elevation (see quote).
Bartholow (1989) found that air temperature above the stream surface was the greatest factor in increasing water temperatures followed in importance by relative humidity and shade, respectively (see graph at left). Bartholow's (1989) work was based on field data from hundreds of locations throughout the West used to develop the SNTEMP stream temperature model. While many previous works considered direct solar radiation to be the dominant mechanism for warming streams (Brown, 1980 as cited in Spence et al., 1998), most of the recent scientific literature considers air temperature over the stream to be the most influential factor (see quote from Essig, 1998 ). Poole and Berman (2000) also recognize the relationship between increasing air flow over the stream and water temperature elevation (see quote).
Halsted (2002) suggested that flow depletion may exacerbate water temperature problems in the Sheepscot River, particularly in late summer and early fall.
 While there are many forest settings in the west with undisturbed riparian conditions, most of the temperate forest of Maine has been harvested at least once and much is still actively managed today. The National Research Council (2004) pointed out that while forests in Maine covered 92% of the landscape before disturbance (1600), that only 53.2% was forested in 1872, but that forests again covered 89.6% of the state by 1995. Although largely forested today, forest condition and health may be much different than forests in a pre-disturbance conditions. Consequently, there is a gap in understanding to what degree historic riparian conditions in old growth forests of Maine may have controlled or buffered water temperatures, and restoring such function may be challenging due to the lack of baseline data. Burgi and Russell (2001) recommend that historians work together with ecologists to reconstruct historic conditions for better understanding of pre-disturbance conditions and the nature of human disturbance and its effects on ecosystems over time. See Riparian Zones and Large Woody Debris Background page for more information.
While there are many forest settings in the west with undisturbed riparian conditions, most of the temperate forest of Maine has been harvested at least once and much is still actively managed today. The National Research Council (2004) pointed out that while forests in Maine covered 92% of the landscape before disturbance (1600), that only 53.2% was forested in 1872, but that forests again covered 89.6% of the state by 1995. Although largely forested today, forest condition and health may be much different than forests in a pre-disturbance conditions. Consequently, there is a gap in understanding to what degree historic riparian conditions in old growth forests of Maine may have controlled or buffered water temperatures, and restoring such function may be challenging due to the lack of baseline data. Burgi and Russell (2001) recommend that historians work together with ecologists to reconstruct historic conditions for better understanding of pre-disturbance conditions and the nature of human disturbance and its effects on ecosystems over time. See Riparian Zones and Large Woody Debris Background page for more information.
Sedimentation of streams may also contribute to elevated water temperatures. Sediment can fill pools and cause the width-to-depth ratio of a stream to increase, which can facilitate heat exchange (Poole and Berman, 2000). Poole and Berman (2000) noted that large wood jams can contribute to stream cooling by forcing more stream flow into shallow ground water, which is called the hyporheic zone. The water drops slightly in temperature before emerging downstream. Whether there are similar benefits from beaver ponds on Maine rivers needs to be explored.
For a synthesis of information on water temperature suitability in the Sheepscot River, see Hypothesis #2: Elevated water temperature is limiting Atlantic salmon production in many reaches and tributaries of the Sheepscot River.
References
Bartholow, J.M. 1989 . Stream temperature investigations: field and analytic methods. Instream flow information paper no. 13. Biological Report 89(17). U.S. Fish and Wildlife Service, Fort Collins, Co. [900 kb]
Baum, E. (1997). Maine Atlantic Salmon: A National Treasure. Hermon, ME:, Atlantic Salmon Unlimited. 224 p.
Burgi, M. and E.W.B. Russell. 2001. Integrative methods to study landscape changes. Land Use Policy 18 (2001) 9-16.
Danie, D.S., J. G. Trial, and J. G. Stanley. 1984. Species profiles: life histories and environmental requirements of coastal fish and invertebrates (North Atlantic) -- Atlantic salmon. U.S. Fish Wildlife Service.. FWVOBS-82/11.22. U.S. Army Corps of Engineers, TR EL-82-4. 19 pp. [500kb]
Dickerson, H. W., and D. I. Dawe. 1995. Icthyophthirius multifiliis and Cryptocaryon irritans (Phylum Ciliophora). Pages 181-227 in P. T. K. Woo, editor. Fish Diseases and Disorders: Protozoan and Metazoan Infections, volume 1. CABI, New York, New York.
DeCola, J.N. 1970. Water quality requirements for Atlantic salmon, USDI. Federal Water Quality Administration, N.E., Region, Boston, Mass. 42 pp.
DeCola, J.N. l975. Atlantic salmon restoration and the question of water quality. Int. Atl. Salmon Found. Spec. Publ. Ser. b:24-28.
Dill, R., C. Fay, M. Gallagher, D. Kircheis, S. Mierzykowski, M. Whiting, and T. Haines. 2002. Water quality issues as potential limiting factors affecting juvenile Atlantic salmon life stages in Maine rivers. Report to Maine Atlantic Salmon Technical Advisory Committee by the Ad Hoc Committee on Water Quality. Atlantic Salmon Commission. Bangor, ME. 28 pp. [162kb]
Duston, J. R.L. Saunders and D.E. Knox. 1991. Effects of increases of freshwater temperatures on loss of smolt characteristics in Atlantic salmon. Can. J. Fish. Aquat. Sci. 48:164-169.
Elliott, J. M. (1991). Tolerance and resistance to thermal stress in juvenile Atlantic salmon, Salmo salar. Freshwater Biology 25, 61–70.
Elliott, J.M. and J.A. Elliott. 1995. The effect of the rate of temperature increase on the critical thermal maximum for parr of Atlantic salmon and brown trout. J. Fish Biol. 47:917-919.
Elliott, J. M. & Hurley, M. A. (1997). A functional model for maximum growth of Atlantic salmon parr, Salmo salar, from two populations in northwest England. Functional Ecology. Vol. 11: 592–603.
Elson, P.F. 1969. High temperature and river ascent by Atlantic salmon. ICES Andromous and Ctatdromous Fish Commission. 12 p.
Essig, D. 1998. The Dilemma of Applying Uniform Temperature Criteria in a Diverse Environment: An Issue Analysis. Idaho Division of Environmental Quality Water Quality Assessment and Standards Bureau, Boise, ID. 34p. [199 kb]
Garside, E.T. 1973. Ultimate upper lethal temperature of Atlantic salmon (Salmo salar). Canadian Journal of Zoology: 51: 898-900.
Guillen, G. 2003. Klamath River fish die-off, September 2002: Causative factors of mortality. Report number AFWO-F-02-03 . U.S. Fish and Wildlife Service, Arcata Fish and Wildlife Office. Arcata, CA. 128 pp.
Halsted, M. 2002. Effects of Stream Flow on the Stream Temperature, E. Coli Concentrations and Dissolved Oxygen Levels in the West Branch of the Sheepscot River. Sheepscot Valley Conservation Association. 15 p. [450kb]
Heggenes, J. 1990. Habitat utilization and prefernces in juvenile Atlantic salmon in streams. Regulated Rivers Research and Management, Vol. 5, p. 341-354.
Hendry, K.and D. Cragg-Hine (2003). Ecology of the Atlantic Salmon. Conserving Natura 2000 Rivers Ecology Series No. 7. English Nature, Peterborough. On line at: http://www.english-nature.org.uk/lifeinukrivers/species/salmon.pdf.
Jensen, A.J., B.O. Johsen, and L. Saksgard. 1989. Temperature requirements in Atlantic salmon (Salmo salar), brown trout (Salmo trutta), and Arctic char (Salvelinus alpinus) from hatching to intitial feeding compared with geographic distribution. Can. J. Fish. Aquat. Sci. 46:786-789.
Jonnson, B. and J. Russ-Hansen. 1985. Water temperature as the primary influence on timing of seaward migration of Atlantic salmon (Salmo salar) smolts. Can. Jour. of Fish. and Aq. Sci. 40:313-321.
LaBar, G.W., J.D. McCleave and S.M. Fired. 1978. Seaward migration of hatchery-reared Atlantic smolts in the Penobscot river estuary, Maine open water movements. J. Cons. Int. Explor. Mer. 38 (2):257-269.
McCullough, D. 1999 . A Review and Synthesis of Effects of Alterations to the Water Temperature Regime on Freshwater Life Stages of Salmonids, with Special Reference to Chinook Salmon. Columbia Intertribal Fisheries Commission, Portland, OR. Prepared for the U.S. Environmental Protection Agency Region 10. Published as EPA 910-R-99-010. [900 kb]
Peterson, R.H., H.C.E. Spinney, and A. Sreedharan. 1977. Development of Atlantic salmon (Salmo salar) eggs and alevins under varied temperature regimes. J. Fish. Res. Board Can. 34:31-43.
Poole, G.C., and C.H. Berman. 2000. Pathways of Human Influence on Water Temperature Dynamics in Stream Channels. U.S. Environmental Protection Agency, Region 10. Seattle, WA. 20 p. [84 kb]
Shepard, S.L. 1991. Report on radio telemetry investigations of Atlantic salmon migration in the Penobscot River. Bangor Hydro-electric Company, Bangor, ME. 35 p.
Shepard, S.L. 1995. Atlantic salmon spawning migrations in the Penobscot River, Maine- Fishways, flows and high temperatures. M.S. Thesis, University of Maine, Orono, ME. 111 p.
Spence, B.C., G.A. Lomnicky, R.M. Hughes and R. P. Novitzki. 1996. An Ecosystem Approach to Salmonid Conservation. Funded jointly by the U.S. EPA, U.S. Fish and Wildlife Service and National Marine Fisheries Service. TR-4501-96-6057. Man Tech Environmental Research Services Corp., Corvallis, OR.
Swansburg, E., G. Chaput, D. Moore, D. Caissie, and N. El-Jabi. 2002. Size variability of juvenile Atlantic salmon: links to environmental conditions. Journal of Fish Biology (2002) 61, 661–683.